三葉虫
三葉虫(さんようちゅう、英語: trilobite トライロバイト、学名: Trilobita)は、古生代にかけて生息した化石節足動物の分類群である。分類学上は三葉虫綱とされ、横で3部に分かれた硬い外骨格を背面にもつ。古生代を代表する海棲動物であり、冒頭のカンブリア紀から繁栄し、終焉のペルム紀末で絶滅した。2万2,000種以上が知られる多様なグループで化石も多産し[2]、示準化石としても重視される。
| 三葉虫 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||
| 保全状況評価 | |||||||||||||||
| 絶滅(化石) | |||||||||||||||
| 地質時代 | |||||||||||||||
| 古生代カンブリア紀第二期 - 第三期境界(約5億2,100万年前) - ペルム紀チャンシンジアン期(約2億5,200万年前) | |||||||||||||||
| 分類 | |||||||||||||||
| |||||||||||||||
| 学名 | |||||||||||||||
| Trilobita Walch, 1771 | |||||||||||||||
| 英名 | |||||||||||||||
| Trilobite |
形態と生理学


三葉虫の大きさは種類や成長段階により様々である。知られるうちで最大のものは元々体長90 cmほどあったと推測される断片化石であり[3]、ほぼ全身の化石に限れば72 cmが最大となるが[4]、小さいものは成体でも1 mm程度である[5]。幼生の化石も発見されており、最も小さな個体は直径0.2 mmほどである。なお、多くの種類の成体は3-10 cmの範囲に収まる[4]。
体は前後に数多くの体節からなり、各体節に一対の付属肢(関節肢)が備わっていたと考えられ、全てが体の下に覆われる。多くの三葉形類と似て、体節は前後に頭部(cephalon)と胴部 (trunk)、胴部は更に胸部(thorax)と尾部(pygidium)と計3つの合体節に分化するが[6]、他の三葉形類と異なり、全身の外骨格のうち背面全体(背板と複眼)と腹面の一部(背板縁辺部の折り返しとハイポストーマ)は炭酸カルシウム質で頑丈に生物鉱化 (biomineralization) されている[7]。これは三葉虫の化石が保存されやすく多産する主因でもある[2]。他のArtiopoda類と同様、頭部に含まれる体節数は少なくとも5節(先節+第1-4体節)があるとされ、それに応じて少なくとも4対の付属肢(触角1対と脚3対)が先節直後の第1-4体節に配置される[6](ハイポストーマを上唇と同様に付属肢由来と考えれば、これは先節由来の付属肢となる)[8]。
背面構造


各体節に由来する数多くの背板 (tergite) のうち、頭部と尾部のものは癒合して一枚板状に、胸部のものは体節ごとに可動に分節した複数の胸節 (thoracic segment) となっている。背板は1対の縦溝 (axial furrow) で縦割りに中央部の軸部 (axis, axial lobe, axial ring 軸環とも[9]) とそれを左右対となって挟む肋部 (pleura, pleural lobe 肋葉とも[9]) となっており、この縦割り三区分 (trilobation) が三葉虫 (trilobite) の名称の由来となっている[10]。軸部はアーチ状に盛り上がり、肋部は比較的平坦であるが、より派生的な系統では肋部の外側が腹側へと傾斜する傾向を持つ。このため、三葉虫が腹側へと丸まった時に胸節肋部の外側域が重なり合い、背板で生体部を覆うこととなる防御姿勢 (enrollment) の構築が可能となり、一部の種ではほぼ球状に丸める[11][12]。
 Symphysopsis の holochroal eye
Symphysopsis の holochroal eye Eldredgeops rana の schizochroal eye
Eldredgeops rana の schizochroal eye



頭部背板(背甲、head shield, cephalic shield)の軸部は頭鞍 (glabella) といい[9]、その左右には癒合した体節の名残である溝 (lateral furrow) が数対並んでいる。通常複眼(側眼)は頭鞍のすぐ左右の肋部領域に1対あるが、形は一般的な腎臓型からタワー状に盛り上がるものや眼柄に突出するものまで多岐し、頭部に対する相対的なサイズもさまざまであり、退化消失した種もさまざまな系統で知られている。複眼は多数の個眼(レンズ)を持ち、その数は最多数千に及ぶ[13]。ほとんどの種では正面と両側面の視覚が優れていたことが明らかにされている[14]。複眼は holochroal eye(個眼が多く密着して一般的な複眼のタイプ、多くの種類が持つ)・schizochroal eye(個眼が少なく角膜に分かれて深く入り込むタイプ、Phacopina類特有)・abathochroal eye(個眼が少なく角膜に分かれるが深く入り込まないタイプ、Eodiscida類特有)の3種類が知られている[13]。肋部には体節構造の名残がほぼ見当たらず、脱皮の割れ目である1対の弱線 (顔線 facial suture[15], submarginal suture) が複眼に沿ってその前縁左右から後方まで走る。これは三葉虫に特有で、他のArtiopoda類と明確に異なる特徴の一つである[7]。

 三葉虫の尾部のサイズのバリエーション
三葉虫の尾部のサイズのバリエーション 大きな尾部をもつ Scutellum
大きな尾部をもつ Scutellum


胸部背板は原則としてほぼ同じ構造の繰り返しで、ごく一部の種類(主にレドリキア目)のみある程度の特化が見られる(途中数節のみ肋部や棘が特に強大・後方数節が特に小さいなど)。尾部背板は通常胸部と同形な体節構造が見られるが、一部はその痕跡が見当たらないほど特化している。尾部は微小なものから頭部より大きいものまで知られており、派生的な系統ほど頭部に対して大きくなる傾向をもつ[2]。
腹面構造

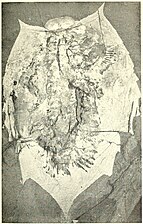
腹面は正中線に1枚のハイポストーマ (hypostome) と1対の脚につき1枚の腹板 (sternite) が並び、背板の縁辺部がわずかに折り返している (doublure)。対になる付属肢は原則として1対の感覚用の触角と複数対の移動用の脚のみを持ち、オレノイデスのみ更に1対の尾毛 (cerci) が確認される。腹板と付属肢対は他の外骨格と比べて硬化していないため化石に残りにくく、ごく一部の種類の良好に保存された化石のみから知られている[6]。
_(17966037580)_(cropped).jpg)




ハイポストーマは現生節足動物の上唇 (labrum) に類する板状構造であり、後方に向けて開いた口を覆い被さる[8]。触角は原則として数多くの小節に分かれた糸状で(アサフェルスは太い棍棒状[16])、ハイポストーマの左右から正面に突き出す。それ以降の腹板は目立たなく、脚はその左右に並び、体節数に応じて頭部に通常3対(一部は直後の胸部との境目まで4対、三葉虫として懐疑的なアグノスタス亜目は5対)、1胸節につき1対、尾部に3対以上を持つ。原則として頭部・胸部・尾部を通じてほぼ同形で、現生節足動物の顎や鋏のように顕著に特化することはなく、大きさや棘の数など細部の連続的な変化のみ確認される[6](オレノイデスのみ著しく特化した脚が確認される[17])。脚は基本的に二叉型 (biramous) であり、主に基部の原節 (protopod, basipod) から腹側に分岐した歩脚型の内肢 (endopod) と外側に分岐し鰭状の外肢 (exopod) で構成される。内肢は先端の爪を含めて7節からなり、棘状の内突起 (endite) を内側にもつ場合が多い。外肢は平たい2節もしくは柱状で数多くの小節に分かれ、後縁に沿って数多くの葉状の構造体 (lamella) が並んでいる。機能的観点から内肢と外肢をそれぞれ単に「歩脚」と「鰓」と呼ばれることが多いが、これは形態のみに立脚する研究を行う古生物学にとって多くの混乱を産み出す要因となっているとのコメントもあり[18]、特に1990年代では外肢の鰓解釈を否定する見解もあった[7]。しかし2020年代の複数の研究により、外肢は解剖学・流体力学的観点から鰓である説が再び有力視されている[19][20][21][22]。顎はないが、原節の内突起は内側に向けて噛み合わせた顎基 (gnathobase) に発達した場合が多い[23][24]。
内部構造

内部構造は消化系のみ明確に知られており、循環系や神経系は不明[25]。ハイポストーマと背甲の間には口から前上方にUターンして曲がり返した食道と前胃が格納され、種類により肥大化した場合がある。それ以降の消化管は一直線に尾部末端の肛門まで達し、多くの場合は頭部から胸部前半にかけて、消化酵素を分泌すると思われる複数対の消化腺 (gut gland, diverticulum) が体節ごとに並んでいる[26][27][28]。
生態

基本的には、海底を這ったり、泳いだりして生活していた底生生物と解釈されている。泥に潜っていた、あるいは浮遊性・遊泳性(外洋性)であったと推測されているものも一部存在し[29]、足跡と考えられる生痕化石 (Cruziana など) も知られている[30]。多くは腐肉食者とされるが、一部の種類は頑強な顎基をもつことにより捕食者とされる[31][24]。貝虫類や棘皮動物、ヒオリテス類などの残骸が含む前胃の内容物も知られている[28]。

成長は他の節足動物と同様、硬い外骨格は成長につれて伸びることができないため、古い外骨格を脱ぎ捨て新しい外骨格に変える脱皮によって行われる。この場合、頭部は表面の弱線に沿って最大5つのパーツに分割され[15]、細部の構造も脱皮ごとに変化が見られる[32]。半増節変態で成長しており、分節を持たない「プロタスピッド」(protaspid)・脱皮ごとに胸節が増える「メラスピッド」(meraspid)・最大の胸節数を保って成長し続ける「ホラスピッド」(holaspid) の3段階に分けられる[6][33]。特にプロタスピッドは三葉虫特有の成長段階であり[34]、種類により成体と似た姿の底生性、もしくは成体と大きく異なる姿で甲殻類のノープリウス幼生のように浮遊性であったとされる[35][36]。なお、知られる卵化石はプロタスピッドより小さいため、それより小型で初期な齢期をもつとも考えられる[37]。
分類
系統位置
| ||||||||||||||||||||||||||||||||||||||||||||||||
| 節足動物における三葉虫の系統位置 |
 ヘルメティア
ヘルメティア



三葉虫はナラオイア、ヘルメティア、シャンダレラなどとも共に三葉形類(Trilobitomorpha)をなし、これは更に光楯類やシドネイアなどと共にArtiopoda類という化石節足動物の一大分類群をなしている[7][38]。その中で三葉虫はConciliterga類(ヘルメティア、サペリオンなど)や Kwanyinaspis との類縁関係が多くの系統解析に支持される[39][40][41][42][43][44][45]。
Artiopoda類は現生節足動物の4つの大きな区分である鋏角類・多足類・甲殻類・六脚類に並ぶ一群としての位置はほぼ認められているが、これらの群との類縁関係については定説がない(鋏角類と大顎類のいずれかに近縁、もしくはそれらより基盤的)[46]。古くは三葉形類から触角が退化消失して前脚が鋏角に変化することで鋏角類が起源し、光楯類やカブトガニ類がその祖先形質を色濃く残した原始的な鋏角類という説があったが[47][48][49]、鋏角は触角に相同であることが多方面な解析に証明される・光楯類は鋏角類的性質をもたないことが解明されるなどの発展に連れて、その説の基準が否定的になった[50][51][52]。
下位分類
古生物学の分類学的辞書ともいえる1953年の『Treatise of Invertebrate Paleontology[53]』においては、三葉虫種全てを三葉虫綱(Class Trilobita)としてまとめている。三葉虫の化石は、ほとんどのものが背側に備わった石灰質の外骨格(背板とハイポストーマ)のみが化石化しているため、分類の定義は背板上の形質に頼らざるを得ない。これは、付属肢などの生体部の形状を重要視して分類同定を行う現生の節足動物群とは根本的に異なるので注意が必要である。つまり不完全な記載分類学的研究ではあるが、その研究の歴史は古く、18世紀後半には三葉虫を節足動物の中の独立したグループとする見解が提出され、また分類学の始祖とされるスウェーデンのカール・フォン・リンネも三葉虫を数種記載している。ただし現今では新種報告の数が著しい減少傾向にあるとされ、主流となる三葉虫研究の方向性の転換も求められているようである。
三葉虫綱内における高次分類群間の類縁関係については、1950年代以降に飛躍的に増加した個体発生過程の情報を用いた研究結果に頼るところが大きい。しかしながら、化石化されない生体部の情報がほぼ完全に欠如しているといったデメリットなどにも起因し、研究者間の見解の一致には未だ遠く、今後も大幅な改訂があるかもしれない。
2010年代現在、22,000種以上の三葉虫が記載されている。昔今の文献によって目は3から11、目不明の科は4から58ほどあるとされる[1]。以下は Fortey 1997 による8目の体系を基に列挙する。

- アグノスタス目 Agnostida
- アグノスタス亜目 Agnostina とエオディスカス亜目 Eodiscina があり、双方とも体サイズが
矮小 ()化しており(通常5-6 mm程度)、前者は2・後者は3の胸節で構成される胸部、そして頭部とよく似た尾部といった特徴を持つ。特にアグノスタス亜目は、頭部に通常認められる複眼を持たない。一方、エオディスカス亜目には abathochroal eye と呼ばれる特殊な形状の複眼が備わる[13]。 - 付属肢はアグノスタス亜目の数種から知られており、他の三葉虫とは大きく異なる特徴を備えている。これは、頭部の脚が5対ある・外肢の附属体は刺毛状であり葉状ではない・前2対の外肢は特に長大・胸部と尾部の脚は外肢が短縮して内肢が櫛状に発達、などが挙げられる[54][55]。この付属肢形態の相違をもとに、アグノスタス目(少なくともアグノスタス亜目)を三葉虫綱に含めない見解もある[1]。化石記録はカンブリア紀からオルドビス紀。
_-_8_mm_1.JPG) Itagnostus interstrictus(アグノスタス目) |  Paradoxides sp.(レドリキア目) |
 Paralejurus sp.(コリネクソカス目) |  Pseudoasaphus praecurrens(アサフス目) |
 Phacops speculator(ファコプス目) |  Cyphaspis sp.(プロエトゥス目) |
 Elrathia kingii(プティコパリア目) |  Ceratarges sp.(リカス目) |
- レドリキア目 Redlichiida
- 平滑な体型をしており、防御姿勢は他の三葉虫ほど丸くない[11]。尾部は通常非常に小さく、胸節の数は多い[6]。顔線を持たないオレネルス亜目 Olenellina と、顔線を有するレドリキア亜目 Redlichiina を含む。化石記録はカンブリア紀のみ。
- コリネクソカス目 Corynexochida
- 肥大した頭鞍を持つグループ。頭部付属肢が特化していたと考える研究者もいる。主にカンブリア紀に繁栄したコリネクソカス亜目 Corynexochina と、オルドビス紀以降に栄えた比較的のっぺりとしたイラエナス亜目 Illaenina、団扇状の尾部を持つティサノペルティス(スクテラム)亜目 Leiostegiina がある。化石記録はカンブリア紀前期からデボン紀中期。
- アサフス目 Asaphida
- のっぺりとした体を持つグループ。幼生期は浮遊生活を送っていたと考えられている。化石記録はオルドビス紀からデボン紀中期。
- ファコプス目 Phacopida
- オルドビス紀以降に非常に栄えたグループ。棘の多い体をもつケイルルス亜目 Cheirurina、特徴的なコブのある頭鞍をもつカリメネ亜目 Calymenina、非常に発達した眼をもつものが多いファコプス亜目 Phacopina が含まれる。眼は個眼が独立して集まった schizochroal eye[13]。化石記録はカンブリア紀末からデボン紀後期。
- プロエトゥス目 Proetida
- 楕円形状の体と曲玉状の眼をもつグループで出現が最も遅く(オルドビス紀)、一番後(ペルム紀末)まで生き残っている。しばしば同種のみがまとまって見つかることも多いため、集団で生活していたとも考えられている。
- プティコパリア目 Ptychopariida
- 比較的特徴のない三葉虫のグループ。最も原始的と考えられているプティコパリア亜目 Ptychopariina、カンブリア紀後期からオルドビス紀に繁栄した胸節が多節化しているオレヌス亜目 Olenina、西洋の竪琴型の頭を持つハルペス亜目 Harpina が含まれる。研究者によっては最後のハルペス類をハルペス目として独立させている。化石記録はカンブリア紀からデボン紀後期。
- リカス目 Lichida
- 装飾の多い頭鞍と、体の縁に棘を持つことの多いグループ。防御姿勢がとれない代わりに棘が発達したとする研究者もいる。以前は別々の目とされていたオドントプレウラ類 Odontopleuridae とダメセラ類 Damesellidae を含む。化石記録はカンブリア紀後期からデボン紀中期。
主な種類
 |  ダルマニテス |
.jpg) テラタスピス |  ワリセロプス |
- アルバーテラ Albertella
- ロッキー山脈などのカンブリア紀層で発見される。全長8 cm。
- アークティヌルス Arctinurus
- ヨーロッパや北アメリカのシルル紀層で発見される。大型で、幅は10 cmくらいある。「三葉虫の王様」とも呼ばれる[56]。
- アサフス(ネオアサフス)・コワレフスキー Asaphus kowalewskii
- ロシア・サンクトペテルブルク近郊のヴォルホフ川流域のオルドビス紀層で発見される。眼柄で突き出した複眼が特徴。全長2.5 cmほど。
- ダルマニテス Dalmanites
- 世界中で発見されている、古生代中頃の最も代表的な三葉虫。幅3 cm。
- ダイフォン Deiphon
- イギリスのシルル紀層で発見される。棘が多い。全長4 cm。
- エオレドリキア Eoredlichia
- 中国のカンブリア紀層で発見される。付属肢と消化系が知られる数少ない種類の一つ。体長12 cm(尾部の長い棘を含むと約20 cm)ほど。
- エルラシア・キンギ Elrathia kingi
- アメリカ合衆国のユタ州のカンブリア紀層で発見される。全長1.6 cmほど。
- オレノイデス Olenoides
- 北アメリカのカンブリア紀層で発見される。最も研究が進んでいる三葉虫の一つで、唯一に特化した脚や尾毛が知られる三葉虫でもある[17]。体長10 cmほど。
- パラドキシデス Paradoxides
- カンブリア紀の大型(全長40 cm前後)の三葉虫。
- フィリップシア Phillipsia
- 日本でも発見された。古生代後期。小型(幅2 cm)。
- テラタスピス Terataspis
- アメリカ合衆国のニューヨーク州やカナダのオンタリオ州で発見されている。70 cmほどになるとみられるリカス目の最大級の三葉虫[57]。
- トリアルトルス Triarthrus
- 北アメリカ、ヨーロッパと中国のオルドビス紀層で発見される。付属肢や消化系がよく知られ、最初に卵が発見された三葉虫でもある[37]。体長5 cmほど。
- ワリセロプス Walliserops
- モロッコのデボン紀層で発見される。トライデント状に突き出した角が特徴で、それをカブトムシのように闘争に使われたという説がある[58]。体長4 cmほど(角を除く)。
出現、繁栄と絶滅
.png)
最古の三葉虫の体化石は古生代カンブリア紀前期(第二期 - 第三期境界、約5億2,100万年前)まで遡り、これは節足動物として知られる中で最古の体の化石記録でもある[59]。かつては化石が見つからないだけで、それより前のエディアカラ紀に起源するではないかという説があったが[60]、前述した化石記録の直前に当たるカンブリア紀内(フォーチュニアンもしくは第二期)に起源する説の方が有力視されている[61][2]。フォーチュニアンの堆積層で見つかった Rusophycus などの生痕化石を三葉虫由来とする説があるが[61]、これは別の節足動物由来の可能性もある[62]。
三葉虫は約2億7,000万年に及ぶ生息時期の中で、古生代前期のカンブリア紀とオルドビス紀に最も繁栄した。カンブリア紀前期からオルドビス紀前期には多くの科が出現と絶滅し、オルドビス紀中期と後期には多様化しつつも科の数が比較的安定していたが、オルドビス紀末期の大量絶滅により約25%の科と60%の属を失い、シルル紀以降は徐々に衰退した。デボン紀前期にはおそらく内海の大量形成に恵まれて、以前ほどではないものの再び多様化したが、デボン紀中期には急速な海水位変化と海洋無酸素事変の影響を受けて大きく衰退した。石炭紀中期以降にはプロエトゥス目の2科のみ生き延びて、最終的にペルム紀末期の大量絶滅に巻き込まれる形で絶滅した[15][62]。
文化
三葉虫は有名な化石生物の一つで、無脊椎動物ではアンモナイトとで双璧をなす。化石を装飾品にすることもあるが、日本ではあまり産出しないため(気仙沼市など東北地方では比較的多い[57])、南アメリカなどから輸入されている。
脚注
参考文献
- リチャード・サウスウッド『生命進化の物語』垂水雄二(訳)、八坂書房、2007年。ISBN 978-4896948875。